



��x����Դˮ̎�����ϏS
(li��n)ϵ�ˣ��O��(j��ng)��
�֙C(j��)��15838253283����̖ͬ(h��o)��
�Ԓ��0371-66557686
�]�䣺1813885391@qq.com
��ַ������ʡ��x�й��I(y��)�@�^(q��)
�����w�S������1�S���ЙC(j��)��Ⱦ��������ÙC(j��)��������Ⱥ�w�Б�(y��ng)�{(di��o)������
�����w�S������1�����a(ch��n)�S���ЙC(j��)��Ⱦ��������ÙC(j��)��������Ⱥ�w�Б�(y��ng)�{(di��o)���������ЙC(j��)��Ⱦ��(����N��ˎ��r(n��ng)ˎ��)��������Ȼϵ�y(t��ng)(������������ˮ��������)�͏Uˮ̎��ϵ�y(t��ng)�У�����ӽY(ji��)��(g��u)��(f��)�s���N�׃���ܽ��Բ���Ը߲����y���オ�⣬�����Ч����ȥ����Ŀǰ�о����y�c(di��n)��
�������\�g(sh��)��̎���ЙC(j��)��Ⱦ�����Ҫ�ֶ�֮һ������ż�Ϲ��ܲ��ϡ�����������﹤ˇ(�煒��-��������늴����)��Ŀǰ�о����c(di��n)��������ڲ�����ȫ�V�������γɏ�(f��)�s���ж����x���a(ch��n)��o(w��)����Ч���ƴ��x�^(gu��)�̡���N��Ⱦ�ﹲ��r(sh��)����Ч�ʵ͵Ȇ�(w��n)�}��
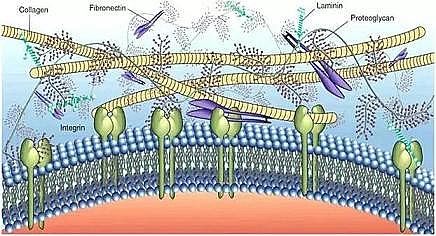
�����������ۺ���(Extracellular polymer subst-ance��EPS)�Ƕ��ǡ������|(zh��)����ֳ�ᡢ���ᡢ֬�|(zh��)���������ӽM�ɵď�(f��)�s�߷��Ӿۺ��Ҳ�������R(sh��)�e��������ȥ���ЙC(j��)��Ⱦ�����Ҫ���ϣ�������H��ˮ�ԡ�����ԡ������ԡ����オ���Լ�����߀ԭ�Ե����ԡ�
����Ŀǰ�������о��༯����EPS��ȡ�y(c��)�������ĸ��M(j��n)�����܈F(tu��n)�M�ɺ����|(zh��)�����䌦(du��)�ЙC(j��)��Ⱦ���ȥ�������ÙC(j��)���о����������ÙC(j��)����Ҫ�漰��������(����)�����W(xu��)����(߀ԭ)�����オ��(����ø��)�����ڷ���ˮƽ���ЙC(j��)��Ⱦ��ȥ���{(di��o)�ؙC(j��)��������ȱ��ϵ�y(t��ng)���о���(b��o)����
����EPS�ϳɼ��ЙC(j��)�オ�������̖(h��o)���Ӻ�Ⱥ�w�Б�(y��ng)�{(di��o)�أ��Ǐ�(qi��ng)����Ⱦ��ȥ������Ҫ�h(hu��n)��(ji��)��ͨ�^(gu��)��������{(di��o)��EPSȥ���ЙC(j��)��Ⱦ����P(gu��n)�I���R(sh��)�e��EPS�c�ЙC(j��)��Ⱦ�����õ��P(gu��n)�I�M�֡�
����EPS���Ӿ��Џ�(f��)�s��׃�ľW(w��ng)�j(lu��)����g�Y(ji��)��(g��u)�����������w����(�R(sh��)�e�ЙC(j��)��Ⱦ��)����Ĥ����(�\(y��n)ݔ�ЙC(j��)��Ⱦ��)����Ⱥ�w�Б�(y��ng)�R(sh��)�eϵ�y(t��ng)؞�����У�����������M(j��n)����Ⱦ���R(sh��)�e������Ⱥ�w�Б�(y��ng)�{(di��o)�ؽ��ⷽ���ϵ�y(t��ng)�Ԉ�(b��o)�����ࡣ
������(du��)�ˣ��P�߾C��������ۼ��w��EPS��(du��)��ͬ�ЙC(j��)���ȥ�����á�Ⱥ�w�Б�(y��ng)�{(di��o)�ؙC(j��)�������c����Ⱥ��Y(ji��)��(g��u)���P(gu��n)ϵ�����ڌ�(du��)���(qi��ng)��ȥ���ЙC(j��)���ṩָ��(d��o)��
����01
��������ۺ���(EPS)�ĽM�ɺ����|(zh��)
����EPS�ɶ���(PS��40%~95%)�������|(zh��)(PN��1%~60%)����ֳ�|(zh��)(HS)������(1%~10%)��֬�|(zh��)(1%~10%)�ȴ�������|(zh��)�M��(�����|(zh��)����?j��n)?sh��)Ӌ(j��))��
��������PS�ɶ�Nͬ����(���Ͼ��ǡ������ǵ�)�ͮ�����(�����}���ʰ����z��)�M�ɡ�����PN������ø�ͽY(ji��)��(g��u)���ף����нY(ji��)��(g��u)���װ�����(x��)���������P(gu��n)����̼ˮ������-�Y(ji��)�ϵ��ס�
�������⣬EPS�е�PN��PS߀��(hu��)���ǾY����(���ġ��ǵ��ס���֬��)����ʽ���ڡ�HS�ɸ�ֳ��(HA)��������(FA)�����|(zh��)�M�ɡ�
����EPS�к�̼���|(zh��)�Ⱥ������|(zh��)�ϳɸ����ٶȸ��죬�纬��������|(zh��) > �����������|(zh��) > ��α̼���|(zh��) > ��֬����̼���|(zh��)�������� > �����|(zh��) > ��ֳ�|(zh��)��֬�|(zh��)�
��������(j��)EPS�c��(x��)��������B�ӳ̶ȼ���ȡ����������ӵ���(n��i)�ӿɷ֞������EPS(SOL-EPS)����ɢ�Y(ji��)����EPS(LB-EPS)���o�ܽY(ji��)����EPS(TB-EPS);����(j��)�Ƿ��γ�ˮ���z��EPS�֞�ˮ���zEPS�ͷ�ˮ���zEPS��
�������У�ˮ���zEPS�ַQ�Y(ji��)��(g��u)EPS(Structural EPS��SEPS)�����ЙC(j��)�M�ɺ��x�������Q�����������|(zh��)��EPS�����H��ˮ�ԡ�����ԡ������ԡ����オ���Լ�����߀ԭ�Ե����ԡ�ԓ���ԛQ������ˮ̎����ˮ�|(zh��)������Ⱦ�オ��̶ȡ���������ij�����Óˮ���ܵȡ�
����02
����EPS���y�����ЙC(j��)���M(j��n)�����:(x��)������Ҫ����
����������������Դ�ЙC(j��)���M(j��n)�뼚(x��)���ͼ�(x��)���g����õ���Ҫͨ�������������ЙC(j��)����н������Ķ�Nøϵ����(x��)���g��̖(h��o)����(d��o)���ֿ���Դ���ͨ�^(gu��)����ˮ���ṩ�I(y��ng)�B(y��ng)���|(zh��)���ЙC(j��)�|(zh��)����Ҫ���ܡ�
����2.1�ЙC(j��)��Ⱦ��İ���(n��i)��������������
��������(j��)�ЙC(j��)��Ⱦ���ڼ�(x��)�����������λ�ã��ЙC(j��)��Ⱦ��ȥ���ɷ֞����N��ʽ��
����(1)����̎������EPS�r���ЙC(j��)�����ø�ڰ���ֱ���������ЙC(j��)��Ⱦ�����ɴ��x��/���a(ch��n)Ʒ���矟��ù(Aspergillusfumigatus)��ȥ����r(sh��)��Ҫ���þ��ǰ���ø(ľ�|(zh��)���^(gu��)������ø)���x��
����(2)�س�̎�������ЙC(j��)��Ⱦ���ڼ�(x��)��ĤEPS��������ͬ�r(sh��)�ڰ���ø�����|(zh��)ø�����|(zh��)ø�������س̽��⡣������ȥ��ʯ�͟N�^(gu��)���У������|(zh��)ø�����|(zh��)ø�Ͱ���ø�����c���オ�⣬ֻ�Dz�ͬ�����ø���Բ�ͬ��
����(3)����(n��i)̎��������Ⱦ���M(j��n)�����(n��i)���ɰ���(n��i)ø���x���ų���������Trametes hirsuta�����ղ���ҡ�����������ͪ��Һ����������ɰ���(n��i)ø(��(x��)��ɫ��c��P450��)������x���ų���(x��)����
����ֵ��ע����ǣ��������|(zh��)����������(����ԴDNA���Ի���)�����EPS�ͼ�(x��)��Ĥ������h(hu��n)��ϵ�y(t��ng)��ȡ����������Ⱦ��Ʌf(xi��)�������ؿ��Ի���(ARG)�@�_(k��i)��(x��)��������(n��i)��ø��(du��)����ԴARGs�Ľ��⣬��EPS�ɝB�����M(j��n)�뼚(x��)�������M(j��n)ARG�ڼ�(x��)���g����������(d��ng)EPS������Ч�r���ЙC(j��)��Ⱦ��r(sh��)����(hu��)���ڼ�(x��)�������L(f��ng)�U(xi��n)��
�������⣬���h(hu��n)���l��׃���r(sh��)������Ɍ�����(n��i)̎���ЙC(j��)��ģʽ�{(di��o)��(ji��)�����̎������Yanxia Zhu�Ȱl(f��)�F(xi��n)�h(hu��n)���иߝ��CO2�ɴ��M(j��n)�g픹�(ji��)����(Arthrospira platensis)����EPS��������������(qi��ng)EPS��������������ʹ��Ĥ�U(ku��)ɢ(����(d��ng)�\(y��n)ݔ)�M(j��n)�뼚(x��)���ı������½���ͬ�r(sh��)������u��ø��������112.7%����K���ӽ����ɰ���(n��i)�D(zhu��n)�������⡣
����2.2�ЙC(j��)��Ⱦ����x�Ķ����T��(d��o)�{(di��o)������
�������:(x��)���ɸ�֪���W(xu��)���|(zh��)��pH���B�����ضȺ�����߀ԭ늄�(sh��)�����h(hu��n)�����ؼ����ݶ�׃�����ЙC(j��)��Ⱦ��ͨ�^(gu��)�Y(ji��)�ϰ��⻯�������w���ס���Ĥ���ף��T��(d��o)�{(di��o)��څ������Ĥ�\(y��n)ݔ���������EPS�ϳ�(���������Ԅ����������)���ЙC(j��)�オ��ø�����D(zhu��n)䛱��_(d��)�����M(j��n)�ЙC(j��)��Ⱦ���������⡣
����(1)څ������ָ��(x��)���m��(y��ng)���W(xu��)���|(zh��)����ݶ������\(y��n)��(d��ng)��׃�������W(xu��)Ч��(y��ng)�����c��(x��)��Ĥ����Ļ��W(xu��)���w����һ�ԽY(ji��)�ϣ���(j��ng)څ����̖(h��o)����(d��o)����(x��)���ʬF(xi��n)څ��ߝ�ȵ������\(y��n)��(d��ng)��͝�ȵ�ؓ(f��)���\(y��n)��(d��ng)�����W(xu��)څ�����w�����ڼ�(x��)���ž��б��ش��ڣ������خ��Ժ�Ĥ��?f��)�Y(ji��)��(g��u)����څ�������飺
����a. �����T��(d��o)����Դ�ЙC(j��)��Ⱦ��μ�(j��)���x�a(ch��n)�����̖(h��o)���Ӿ����T��(d��o)���W(xu��)څ�����ܶ༚(x��)��ֻ��(du��)�ɽ���������څ���ԣ���نΰ���Pseudomonas sp. JHNֻ��(du��)�ܴ��x��4-��-2-�����Ӿ���څ���ԣ���(du��)���ܴ��x�ĵ���](m��i)��څ���ԡ�����(li��n)�������Pseudomonas sp. B4��(j��ng)���g���x�a(ch��n)��(li��n)�������T��(d��o)���B(y��ng)��(du��)�������4-�ȴ����������څ�����á�
����b. EPS�ϳɻ��ë���ơ���(x��)��څ���\(y��n)��(d��ng)�����ɱ�ë����;�෴����(d��ng)����Ҫ�p���\(y��n)��(d��ng)�r(sh��)���t��(qi��ng)���ϳ�EPS�������d�w�����ճ����ԓڅ���^(gu��)��ͨ���ɲ�ͬ�Ļ��W(xu��)�Б�(y��ng);��(chemosensory path-way)��ɣ���ճ���Myxococcus xanthusͨ�^(gu��)Dif;�����Ƽ�(x��)������ǵĮa(ch��n)����֬�|(zh��)��څ���ԡ�ˮ����(x��)���Թȿݲ���(Burkholderia glumae)��QS(Ⱥ�w�Б�(y��ng))����qsmR���{(di��o)�ر�ë�������ı��_(d��)����y����(Vibrio cholerae)��CAI-1Ⱥ�w�Б�(y��ng)ϵ�y(t��ng)�ܾ�Ⱥڅ������״ر��_(d��)���ƣ��䲻ͬ���L(zh��ng)�ڵļ�(x��)���ΑB(t��i)��EPS���\(y��n)��(d��ng)��c-di-GMP(�h(hu��n)���B(ni��o)����)�D(zhu��n)��{(di��o)�ء�
����c. ���M(j��n)�������úͽ��⡣��(x��)��ͨ�^(gu��)���W(xu��)څ���^(gu��)�̴��M(j��n)��(du��)����ˮ�Ժ͵͵����������ЙC(j��)��Ⱦ��Ľ��⡣���^�����نΰ���G7��څ���Ժͷ�څ���Ծ���l(f��)�F(xi��n)���ЙC(j��)��Ⱦ���ڷ�ˮ��Һ�w(NAPL)�Еr(sh��)��څ����(hu��)�������Ľ�����������öȡ�
����d. �p����(Biphasic chemotaxis)����(x��)���S���h(hu��n)�����ЙC(j��)��Ⱦ����׃���������ؓ(f��)���\(y��n)��(d��ng)���������� < 1 mmol/L�r(sh��)���c�U��E. coli�Hؓ(f��)���(q��)��(d��ng)(�(q��)�܄�����(y��ng))��������≥1 mmol/L�r(sh��)����ؓ(f��)�������(q��)��(d��ng)(����������(y��ng))�ĕr(sh��)�g��ه�Է��D(zhu��n)��
����(2)��Ĥ�\(y��n)ݔ����Ĥ������؞������(g��)��֬�p�ӵă�(n��i)��Ĥ���ף���ɶ˷քe��¶�ڼ�(x��)��Ĥ���ⲿ�̓�(n��i)�����ڼ�(x��)���c�h(hu��n)��֮�g���Q���|(zh��)����������Ϣ���S�������ڽ����y�����ЙC(j��)��Ⱦ��r(sh��)��(hu��)��QS�T��(d��o)�a(ch��n)����ͬ���黯���ͱ�����Ԅ�(����֬��֬�ġ��x��֬�|(zh��)������֬�|(zh��)�;ۺ����������Ԅ�)�����б�����Ԅ�Ҳ�ɽ�����Һ���揈�����黯���ã����Ƽ�(x��)���\(y��n)��(d��ng)�ԺͿ�Ĥ���ױ��_(d��)��
������(x��)����Ĥ���������(du��)PAHs(��h(hu��n)���N)�D(zhu��n)�\(y��n)�^(gu��)���е��������P(gu��n)��Ҫ����ԭ�����ڵ��͵Ŀ�Ĥ���׃�(n��i)����Ҫ����ˮ��β-�ۯB�M�ɣ����H��α-������Ҫ�ֲ����������α- �����ɴ��M(j��n)������Ԅ��ۼ��ͼ�(x��)��Ĥ����֬�p���ں��γɻ�ϵ��z������׃��(x��)��Ĥ������(j��)�Y(ji��)��(g��u)�͘�(g��u)��β-�ۯB���M(j��n)�˱�����Ԅ�������֬�p�ӣ��Լ���PAHs��(x��)����(n��i)���D(zhu��n)�\(y��n)������(qi��ng)��(x��)���c����Ĥ��(du��)��Ⱦ����������c���❓�ܡ�
����(3)�D(zhu��n)��{(di��o)�ء��D(zhu��n)��{(di��o)�����{(di��o)�ؼ�(x��)��������_(d��)���m��(y��ng)����׃���h(hu��n)���l���Ļ����C(j��)��֮һ������Ȼ���У�׃��(g��u)�D(zhu��n)�����aTF(allosteric transcription factor)���R(sh��)�e�����ء�һ��(j��)�����(j��)���x�a(ch��n)�Ⱥ�w�Б�(y��ng)��̖(h��o)���ӻ���Դ�ЙC(j��)��Ⱦ��c��(x��)���Ľ�(j��ng)��ֽ���x�ٿv���෴�������(sh��)������Դ�ЙC(j��)���������õĵ������D(zhu��n)䛼������ӵ���ʽ���F(xi��n)(CatM���⣬����һ�N��������)�������������܆�һ�����|(zh��)NahR���Ƶġ������D(zhu��n)�����(TFs)�錧(d��o)�������D(zhu��n)�߀�ܼ�(x��)��������r�Q����ȫ���{(di��o)�������{(di��o)��(ji��)��
����03
����EPS�c�ЙC(j��)��Ⱦ���g�����ÙC(j��)��
����EPSȥ���ЙC(j��)��Ⱦ��C(j��)��������������(����������)�����W(xu��)����(߀ԭ��늻��W(xu��)߀ԭ)����������(����ø����)���ш�(b��o)�����ЙC(j��)��Ⱦ����Ҫ�����ڷ���N(�š���)��Ⱦ�ϡ��ױ����{(l��n)(TB)���A���{(l��n)54(BB54)�Ϳ�����(�ǰ�����ء��ĭh(hu��n)��)���������ȡ�
�������ִ�������Ⱦ�P��EPSȥ���C(j��)�Ƽ��z�y(c��)�ֶ�Ҋ(ji��n)�� 1��
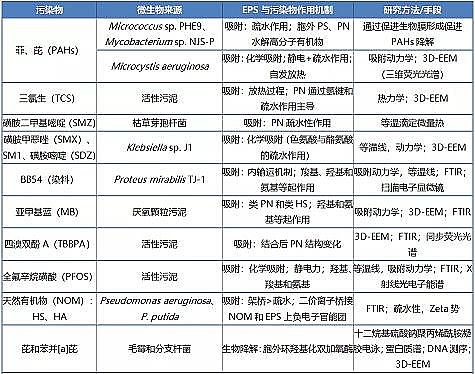
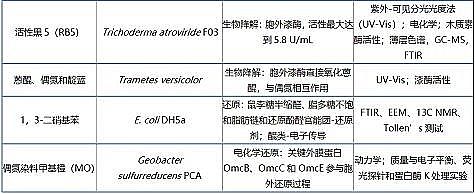
������ 1 EPSȥ���ЙC(j��)��C(j��)�Ƽ��z�y(c��)�ֶ�
����3.1EPS�c�ЙC(j��)��Ⱦ��֮�g����������(����/��������)
����(1)�������á�ָˮ��Uˮ���ЙC(j��)��Ⱦ������EPS���������еķ����^(gu��)�̡�ԓ�������棬�c��������λ�o(w��)�P(gu��n)��ֻ�c�ضȻ��ЙC(j��)���ܽ�����P(gu��n)����EPS����֬(�ɟo(w��)ɫ��PS1��ѿ�ߗU��SLDB1���~�G�نΰ���S5�a(ch��n)��)��֬��(���n�חU���a(ch��n)��)���������Ԅ����ԏ�(qi��ng)��PAHs���������ã����M(j��n)PAHs�����:(x��)���Ă��|(zh��)���������ǺͰ����ǽM�ɵİ�����Ǿ��������黯����(��ꎜ��c�U��TUጷ�)���������ʮ�����������öȣ����M(j��n)�併�⡣
����(2)�������á�����EPS���д���늺ɡ��O�Ի��F(tu��n)����ˮ�^(q��)���ЙC(j��)��Ⱦ���ṩ������������λ�c(di��n)�����漰���I�������A�����o�����ú���ˮ�����4�N������������
����a. ��ˮ����á���ˮ��������ˮ���|(zh��)(��ˮ���܈F(tu��n))֮�g������ˮ�������Ј�(b��o)��(d��o)�Q��������������ˮ���|(zh��)����Ě��w(ˮ������ܽ��Ԛ��w)�(li��n)ͨ�γɵļ{�ך��ݘ����𡣰���PN��ɫ������Ұ����Ǿ��Џ�(qi��ng)��ˮ��(c��)朵���ˮ������(������h(hu��n))�����ṩ��ˮ����λ�c(di��n)��ͨ�^(gu��)���I(���������u��)����ˮ����������Ⱦ��;HS����ؓ(f��)늵�����������������ˮ�^(q��)���̼�Ǽܣ���ͨ�^(gu��)��ˮ���ýY(ji��)����Ⱦ�Ŀ��(bi��o)��Ⱦ��ĽY(ji��)��(g��u)��Ɍ�(d��o)����Ⱦ����EPS�Ͼ߲�ͬ���H����������Ч�ʡ���EPS�е�ɫ���ᡢ�Ұ����c�������ͨ�^(gu��)��ˮ����ýY(ji��)���γ�EPS-��������(f��)����ǰ��א���(SMX)��(c��)�����gλ��С����ռ��(j��)��������λ��EPS����Ч�ʸߣ����ǰ����(SDZ)��ˮ���ò���ӽ�EPS����λ�c(di��n)���������Ч�ʵ͡�
����b. �o����á��o������ǎ���늵��ЙC(j��)��Ⱦ���c��ؓ(f��)늵�EPS֮�g�l(f��)������(y��ng)����Ч�C(j��)�ơ��o�������pH���x�ӏ�(qi��ng)�ȵ�Ӱ푣����cEPS�x�ӻ����܈F(tu��n)(���Ȼ���������F(tu��n)�������ͷ�����܈F(tu��n))�ĺ�������Ⱦ�������P(gu��n)����ױ����{(l��n)(TB)��ͨ�^(gu��)�o�����������EPS�x�ӻ����܈F(tu��n)�ϣ���pH��11.0�r(sh��)����EPS��(du��)TB�ĽY(ji��)��������pH��7.0�r(sh��)���@����?y��n)�?d��ng)pH��7.0�r(sh��)���H�Ȼ���������F(tu��n)����x;pH��11.0�r(sh��)���Ȼ����������������������F(tu��n)�ֶ�����x����EPS�����ڴ�pH�¿�ˮ������ǵ����ж����I�����ӎ�늹��܈F(tu��n)��
����c. ���I�����I���ؓ(f��)�Դ돽С��ԭ��(��O��N��F)֮�g��Hԭ�Ӟ�ý���γ���ƃr(ji��)�I�ĽY(ji��)��(g��u)���߷����ԡ������ԺͼӺ��ԡ���Ⱦ���cEPS֮�g��ͨ�^(gu��)���I�Y(ji��)�ϣ����ĭh(hu��n)���c�����w������EPS�ĵ����|(zh��)��ֳ���ͨ�^(gu��)���I�ͷ����A���γɏ�(f��)���
����ͨ��EPS�c��Ⱦ��֮�g��N�����ͬ�r(sh��)���ڣ������S���l��(pH���x�ӏ�(qi��ng)�ȡ����܈F(tu��n)�NͽY(ji��)��(g��u))׃�������ÙC(j��)��Ҳ�l(f��)��׃����EPS��(qi��ng)��ˮ�ԡ�����λ�c(di��n)�༰��������(qi��ng)ʹ��ȥ���ʸ��ߡ�EPS�c�ЙC(j��)��֮�g�������Y(ji��)�ϙC(j��)�����������ԺͲ�ԣ�����ҳ��@Щ��ā�(l��i)Դ�Ķ�ͨ�^(gu��)����EPS�c�ЙC(j��)�����õ��P(gu��n)�I�M�ֵ����Ɂ�(l��i)��(sh��)�F(xi��n)����EPSȥ���ЙC(j��)���Ч���dž�(w��n)�}���P(gu��n)�I��
����3.2EPS�c�ЙC(j��)��Ⱦ��֮�g�Ļ��W(xu��)����(߀ԭ��늻��W(xu��)߀ԭ)
����EPS����PS�İ�sȩ߀ԭ�Y(ji��)��(g��u)��߀ԭ����(��(x��)��ɫ��c���F��������)��HS������������Ⱦ�������߀ԭ���Ի���ӂ���(d��o)�Եijɷ֡���N��(x��)��EPS����߀ԭ�������N(NAC)���u���N��ż��Ⱦ�ϵȺ���(qi��ng)����ӹ��܈F(tu��n)(���������u����)���ЙC(j��)��Ⱦ�
����(1)늻��W(xu��)���Ծ�EPS߀ԭ���á�늻��W(xu��)���Ծ�EPS����늻��W(xu��)���|(zh��)���䌧(d��o)����|(zh��)��ͨ�^(gu��)늻��W(xu��)߀ԭ���Å��c�ЙC(j��)��Ⱦ�������D(zhu��n)�ơ������늻��W(xu��)���Ծ�Shewanella oneidensis MR-1��Bacillus sp. WS-XY1��Pichia stipites��EPS����늻��W(xu��)���ԣ����w���F(xi��n)�飺
����(a)��(x��)����EPS���w�r(sh��)��ֱ�Ӱ�������D(zhu��n)��(Extracellular electron transfer��EET)�p��;
����(b)EPS�Ѓ�(ch��)���늻��W(xu��)�������|(zh��)(�S�غͼ�(x��)��ɫ��c)����������D(zhu��n)�ƽ��|(zh��)�S�ּ�(x��)���c��ӹ��w/���w�g϶�иߝ����Ӵ�������g��EET��ӂ��f�ӏ�(qi��ng);
����(c)��(d��ng)EPS����ȡ��(hu��)��(d��o)�¼�(x��)�����浰��ʧ���ʧȥEET���á�Mtr����ͨ·���ЙC(j��)��Ⱦ����������D(zhu��n)������Ҫ��ӂ��f;������ϣ���Ͼ�MR-1��ͨ�^(gu��)��(x��)��ɫ��c(MtrC/OmcA)ֱ��߀ԭ��ͨ�^(gu��)������ӽ��w(�S��)�g��߀ԭ��(du��)���ӾGȾ��Óɫ;��ֳ��HS���˿���������D(zhu��n)��˲�r(sh��)���|(zh��)��Ҳ������ֱ����ӹ��w�����cż��Ⱦ�ϡ��u������������廯�������Ⱦ���߀ԭ�^(gu��)�̣��������HS������(hu��)����늻��W(xu��)����(y��ng);�������M(j��n)���������öȺͽ�������;�����|(zh��)��ͬ�r(sh��)�䮔(d��ng)������w����ӹ��w�����c����߀ԭ���ȱ���Ó�u�^(gu��)�̺Ͱ�������D(zhu��n)���^(gu��)�̡�
����(2)��늻��W(xu��)���Ծ�EPS߀ԭ���á���늻��W(xu��)���Ե������ڛ](m��i)����Դ��ӹ��w����r�£�������EPS��߀ԭ�Գɷ�ͨ�^(gu��)��������D(zhu��n)�ƙC(j��)��߀ԭ�������B(t��i)��Ⱦ��(��NAC���t���}�������})��
����Fuxing Kang�Ȱl(f��)�F(xi��n)����N�����B(y��ng)����(���c�U�����S��ԭëƽ�����ᄾƽ�ĸ)����Ȼ����Ĥ����ȡ��EPS�܌�1��3-��������(1��3-DNB)߀ԭ��3-�u������������3-�������������V����1��3-������������(y��ng)ǰ���EPS���Y(ji��)��Tollen�z�(y��n)������EPS�������ǵİ�sȩ����֬���ǵIJ��֬��机ͷӻ�������߀ԭ��������������Ӵ������á�Xinwei Zhou�����о���늻��Լ�(x��)��EPS߀ԭNAC�C(j��)�ƕr(sh��)�l(f��)�F(xi��n)���o(w��)��Դ��ӹ��w�r(sh��)��(x��)������EPS��߀ԭ�ǵİ�sȩ���F(tu��n)(��Ҫ��ӹ��w)�Ԯa(ch��n)����ӣ����ͨ�^(gu��)����D(zhu��n)�ƽ��|(zh��)(�S�غ���)���f����(x��)���⼚(x��)��ɫ�أ��ق��f����(x��)����(n��i)����߀ԭ���Ե���߀ԭNAC��
����3.3EPS�c�ЙC(j��)��Ⱦ��֮�g����������(ø���オ��)
����EPS�а���ø��Ҫ��������ˮ��ø������߀ԭø������Կ�Ӱ��ЙC(j��)��Ⱦ��(Ⱦ�ϡ���h(hu��n)���N������(li��n)�����ЙC(j��)���)�������ʡ�
����(1)����ˮ��ø����Ҫ����̼ˮ������ˮ��ø(�����ø���w�S��ø)������ˮ��ø����ø������ø��ֲ��ø��֬��ø�ȣ���ˮ���ǻ��������������܈F(tu��n)�����������EPS����ȡ��ø�ɴ�ˮ��β-�������_(d��)��ù��;a(ch��n)�A�نΰ����Ľ��|(zh��)ø(PpCutA)�ͺ���نΰ�������ø(PpelaLip)��ˮ����N��ͬ�Y(ji��)��(g��u)�������Ⱦ�
����(2)��������߀ԭø����Ҫ�������خ����^(gu��)������ø����P450����ø���^(gu��)������ø��ľ�|(zh��)���^(gu��)������ø(LiP)���i�^(gu��)������ø(MnP)�ȡ�����a(ch��n)���ķ��خ����^(gu��)����ø(P450�μ���ø��Ѫ�t���^(gu��)������ø)�Ĵ������c����(n��i)�μ���ø�����������������EPA��(y��u)���ЙC(j��)��Ⱦ�
����Chunyun Jia�Ȍ����z�F(tu��n)(Zoogloea sp.)��ȡ��EPS���ڽ����������ţ��Y(ji��)������EPS�Йz�y(c��)������ø���������ø���^(gu��)������ø�����Էքe��(0.41±0.04)U/L��(59.67±3.51)U/L��(55.05±1.13)U/L���Ž�����������Ҫ�����á������(y��ng)�l(f��)�F(xi��n)����EPS��֧�U����(du��)�źͱ���[a]�Ž����ʷքe������9%��6%��ëù����(du��)�š�����[a]�ŵĽ����ʷքe������9%��5%�������|(zh��)�V�l(f��)�F(xi��n)EPS�д���PAHs�P(gu��n)�I����ø�������ጷŰ�����ø(�����_(d��)90~100 U/L)�ֽ�Ⱦ�ϣ���ͬȾ�Ϸֽ�C(j��)�Ʋ�ͬ������ø��ֱ����������������ø�cż���͵��{(l��n)�������С���Ӵ��x���{(di��o)��(ji��)��ľ���������LiP�ɽ������90%�ķ����Ⱦ�
����EPS�c�ЙC(j��)��֮�g�����������W(xu��)���������ø���ؕ�I(xi��n)������(b��o)���^�١������о��l(f��)�F(xi��n)ճ������En-sifer adhaerenȥ������(li��n)��(PCB)�r(sh��)�����������������Ҫ���ã����S�����B(y��ng)�r(sh��)�g���L(zh��ng)���オ������Խ��(l��i)Խ��Ҫ�������オ���cLB-EPS�ж��Ǻ�ø���Գ������P(gu��n)��δ��(l��i)�Ƿ�ɲ���ģʽ���������EPS�����P(gu��n)���ýM�֣��������c��Ⱦ�オ�����P(gu��n)�����P(gu��n)ϵ������(zh��n)�_��(g��u)���ЙC(j��)����xģ�����д��о���
����04
����Ⱥ�w�Б�(y��ng)�{(di��o)���ЙC(j��)��Ⱦ��ȥ���C(j��)��
����Ⱥ�w�Б�(y��ng)(QS)�Ǽ�(x��)���g�Ľ����^(gu��)�̣�����(x��)����ͨ�^(gu��)���ںБ�(y��ng)�ض��Ļ��W(xu��)���|(zh��)(��̖(h��o)����)���{(di��o)��(ji��)������_(d��)��(l��i)�{(di��o)��Ⱥ�w�ܶȡ�QS�����{(di��o)��(ji��)����Ĥ���γɡ����������Ԅ��Įa(ch��n)����������ǵĺϳɡ�ˮƽ�����D(zhu��n)�ơ����x������_(d��)���\(y��n)��(d��ng)�Ժ�څ���Ե����P(gu��n)�Ļ�����_(d��)��
����4.1�{(di��o)���ЙC(j��)��Ⱦ�オ�����
�����S�༚(x��)��ͬ�r(sh��)����QS�������߽z�����(n��i)��/���T��(d��o)��(AHL/AI)�ϳ�ø����ϵ�y(t��ng)�ͷ��N���⡲����h(hu��n)�u���p����ø(RHD)����ϵ�y(t��ng)��
�������~�G�نΰ���CGMCC1.860����rhl QSϵ�y(t��ng)����a(ch��n)����HHL(������-L-�߽z�����(n��i)��)��BHL(������-L-�߽z�����(n��i)��)���c�{(di��o)�ر��ӽ��⡣�M(j��n)һ���о��l(f��)�F(xi��n)��CGMCC1.860 rhl QSϵ�y(t��ng)���{(di��o)��nahH�����a�����2��3-�p����ø(C23O)�Ļ����nahR(�D(zhu��n)䛼�������)�����M(j��n)���������gλ�ѽ�;��������N���オ�⡣�~�G�نΰ���PAO��LasI-��RhlI-����3OC12-HSL��C4-HSL�a(ch��n)��������̖(h��o)���w��L(zh��ng)uxR-(LasR��RhlR��QscR)��LuxRͬԴ��ه��AHLs(C10-HSL)����ant�ٿv�ӵ��D(zhu��n)䛼����ӣ�ant�ٿv�Ӿ��a�������������}�����������ѭ�h(hu��n)���g�w��ø��AHLsӰ���Ⱦ�オ����������x��
�������⣬���ʰ����U��ERW19��novR1/novI1��novR2/novI2 QSϵ�y(t��ng)������ه�@�ɂ�(g��)QSϵ�y(t��ng)�������嶾��(MC)�������(mlrA)�D(zhu��n)䛏Ķ����{(di��o)��MC���⡣
����4.2�{(di��o)��EPS�ɷִ��M(j��n)�ЙC(j��)�オ��
����Ⱥ�w�Б�(y��ng)��̖(h��o)����ͨ�^(gu��)�{(di��o)��EPS�a(ch��n)���g�Ӵ��M(j��n)�ЙC(j��)��Ⱦ�オ��(Ҋ(ji��n)�� 2)������̖(h��o)�����{(di��o)��EPS�ĽM��(���⵰�ס�������ǡ�������֬��)׃�����M(j��n)��Ⱦ�オ�⡣
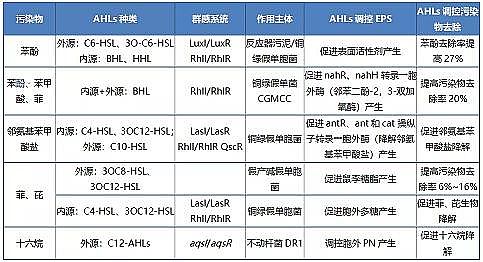
������ 2 �{(di��o)���ЙC(j��)��Ⱦ��ȥ����EPS�ϳɵ�AHLs������r
����(1)���M(j��n)����Ĥ�γɺ�PS/PN�ϳɡ��c�Ҹ���(x��)����ȣ�����Ĥ�е�����ɵֿ�����ж����W(xu��)�h(hu��n)����pH����(d��ng)�ȵIJ���Ӱ푣�����Ⱥ�w�Б�(y��ng)�ʹ��x�a(ch��n)���˼������ЙC(j��)�オ�⡣
����Xi Tang���\(y��n)�ô��x�M�W(xu��)�l(f��)�F(xi��n)������������AHLs��Ҫͨ�^(gu��)����Ala(������)��Val(�i����)��Glu(�Ȱ���)��Asp(�춬����)��Leu(������)�Ȱ����Ậ����(l��i)���M(j��n)��(x��)���⵰�����ɣ��Ķ����M(j��n)����Ĥ/EPS�ĺϳ��ԏ�(qi��ng)��ʮ���顢���������ױ����ЙC(j��)���ȥ����Yue Gu���������(Lactobacillus plantarum)�����Ӳ�ͬ��ȵ�AI-2���M(j��n)��EPS�a(ch��n)�����@����?y��n)�lamC(���a����ˮ��ø����)���a�ĵ����|(zh��)��ˮ����N���ǣ���(du��)β-(1��3)-��β-(1��6)-�B�ӵ��Ͼ��ǵ�����F(xi��n)��ø���ԣ�����(du��)β-(1��4)-�B�ӵ��Ͼ��Ǻ�ľ���ǵ�����F(xi��n)����(n��i)��ø���ԣ�ftsH(���aĤ������ø����)���P(gu��n)��FtsH�����ձ������Ĥ�ϣ��ɽ���Ĥ����;AI-2������lamC��ftsH�Ļ�����_(d��)�Դ��M(j��n)EPS�a(ch��n)���c����Ĥ���γɡ�
����(2)���M(j��n)����ø���_(d��)��ˮ��ø��ˮ���(f��)�s�ЙC(j��)�|(zh��)�͠I(y��ng)�B(y��ng)��V����������Ҫ���ã���Pantoea ana-natis B9�a(ch��n)����6�NAHLs(C4-HSL��3OC6-HSL��C6-HSL��C10-HSL��C12-HSL��C14-HSL)��������(qi��ng)����ˮ��ø(�A������ø)�Ļ���;�՚W���Ͼ�(Erwinia chrysanthemi)����ExpI/ExpRȺ��ϵ�y(t��ng)����a(ch��n)����3OC6-HSL�ɿ��Ƶ���ø�ϳ�;��ˮ��ΰ�������AhyI/AhyRϵ�y(t��ng)����a(ch��n)����C4-HSL�ɿ��ƽz���ᵰ��ø�ͽ��ٵ���ø���a(ch��n)��
����(3)���M(j��n)������Ԅ��ϳɡ�������Ԅ��Ǵ��M(j��n)��(x��)�����������պͿ�Ĥ�\(y��n)ݔ�ЙC(j��)����P(gu��n)�I���|(zh��)����AHLs��ͨ�^(gu��)Ӱ푼�(x��)��������ԡ���ˮ�Ժ��黯����������������ͽ������ܡ����~�G�نΰ�����ͨ�^(gu��)�a(ch��n)������������֬(��֬)������ˮ�Ի�����������ʣ���(qi��ng)����ˎ��ʳƷ�����۵��ИI(y��)�������ޏ�(f��)��
������(d��ng)��ԴC4-HSL���ڕr(sh��)���~�G�نΰ���N6P6��(x��)��������ˮ�Ԟ�51.33%(�o(w��)AHLs�ĿհM������35.33%)���ƵĽ�����������93.3%(���հM�H��85.6%)���~�G�نΰ����{(di��o)��(ji��)ԓ������֬�ϳɵ���ҪQSϵ�y(t��ng)��las��rhl��pqsϵ�y(t��ng)�����w�{(di��o)���^(gu��)�̞飺���ȣ�RhlI�ϳ�ø�a(ch��n)����̖(h��o)����C4-HSL;֮��C4-HSL�cRhlR�Y(ji��)�ϣ�����rhlAB(������֬�ϳ�ø)�����D(zhu��n)䛣��Ķ��ڼ�(x��)����(w��n)�����L(zh��ng)�A�Ά���(d��ng)������֬������ϳɡ�
�������⣬QS�{(di��o)��(ji��)�ЙC(j��)�オ���^(gu��)�̺͙C(j��)�����о��N�خ��Ժ͏�(f��)�s�ԡ���Ⱥ�w�Б�(y��ng)��Novosphingobium pentaromativorans US6-1����novI/novR-QSϵ�y(t��ng)��novI/novRͻ׃�����(sh��)PAH����ø(��ʼøRHD��4-�u����������-3-�μ���ø��ˮ��ȩÓ��ø)�����S�����{(di��o)����EPS�a(ch��n)�����{(di��o)����ȥ����������
�������w��(l��i)�v��AHLs�{(di��o)��EPS�a(ch��n)�����M(j��n)��Ⱦ�オ������ÙC(j��)���Բ����ƣ�Ŀǰ�H�����˽�AHLs��ͨ�^(gu��)���M(j��n)������֬������ø�Įa(ch��n)�������M(j��n)����Ⱦ�オ�⣬��EPS�����M���Ƿ���΅��c��Ⱦ�����オ���Բ�������δ��(l��i)�д����÷�������W(xu��)���g(sh��)(����M�W(xu��)�����M�W(xu��)���ǽM�W(xu��)�����x�M�W(xu��))�M(j��n)һ����ʾEPS�ľ��w�M��(�����ᡢ����)���书�ܣ���Uˮ����̎����AHL�{(di��o)���ЙC(j��)��Ⱦ�オ���ṩ��Փָ��(d��o)��
����4.3�{(di��o)�������\(y��n)��(d��ng)�cڅ��
����QS���{(di��o)��(ji��)��(x��)���\(y��n)��(d��ng)�ԣ��˹��ܞ齵����Ⱦ������ü�(x��)���M(j��n)�������ޏ�(f��)�ṩ�˸���ķ�������rhlI���{(di��o)��(ji��)�~�G�نΰ����ļ�Ⱥ�\(y��n)��(d��ng)��QS����qsmR���{(di��o)��Burkholderia glumae��ë�������ı��_(d��)��E. coli K-12���F(xi��n)��څ��AI-2��څ���\(y��n)��(d��ng)�����������о����c�U���ܶȸБ�(y��ng)�{(di��o)��(ji��)��C(Quorum sen-sing E.coli regulator C��QseC)���������ֲ���Ⱦ�е����Õr(sh��)�l(f��)�F(xi��n)�����c�U��QseC����ͨ�^(gu��)�{(di��o)�ؼ�(x��)�����\(y��n)��(d��ng)���{(di��o)��(ji��)��(x��)������Ĥ�γɡ���(d��ng)��(x��)�����X(ju��)���d�w���棬QS����rhl�_(k��i)ʼ���_(d��)�{(di��o)�خa(ch��n)��EPS�ͱ�����Ԅ����Դ��M(j��n)PAH�ܽ���M(j��n)�뼚(x��)������ͨ�^(gu��)��ë��������څ�����γ�����Ĥ�ͼ���PAH���⡣
����4.4�{(di��o)������ˮƽ�����D(zhu��n)��
������(x��)��Ⱥ���g��(j��ng)���l(f��)��ˮƽ�����D(zhu��n)��(HGT)��(x��)���g�z�����|(zh��)�M���D(zhu��n)�ƣ���Ⱥ�w�Б�(y��ng)��ͨ�^(gu��)�ɷN��ʽHGT�{(di��o)���ЙC(j��)�オ�⣺
����(1)��(d��ng)�|(zh��)�����D(zhu��n)�����ϔy�����a������Դ��Ⱦ��ķֽ���x����r(sh��)����(x��)��ͨ�^(gu��)QS�{(di��o)��DNAጷź��D(zhu��n)���ԏ�(qi��ng)����Ⱦ�オ�⡣QS����rhl��(du��)����Ĥ�л����D(zhu��n)������Ҫ���ã�rhlRؓ(f��)؟(z��)����DNAጷţ�����comX����ã�comX�tؓ(f��)؟(z��)��(x��)����(du��)��(x��)����DNA�����պ��D(zhu��n)����
����(2)AHLsλ�ڿ��Ƅ�(d��ng)�z��Ԫ��(Mobile genetic elements��MGE)�����|(zh��)������(x��)����DNA���ɾ��w�ȡ������P(gu��n)���������ڲ�ͬ��(x��)���g�D(zhu��n)�ơ�����������HGT������(qi��ng)����ĤȺ���е����オ�⣬����Ĥ�м�(x��)���|(zh��)��Ⱦɫ�w�D(zhu��n)��Ч�ʱ����x�B(t��i)��(x��)�����D(zhu��n)���ʸ�10~600����
����4.5�{(di��o)��(y��u)���������L(zh��ng)��Ⱥ��Y(ji��)��(g��u)
����4.5.1 �����wϵ
���������wϵ��QSϵ�y(t��ng)�ܿ������h(hu��n)���l������Դ�ЙC(j��)��������g���x�a(ch��n)���(du��)��Ⱦ�オ�������_(d��)�M(j��n)�����{(di��o)�ػ�ؓ(f��)�{(di��o)�ء���Դ�ЙC(j��)��Ⱦ���{�ȕr(sh��)����(x��)����������ԃ�(y��u)���ڴ��x���ԣ����w�{(di��o)�ط�ʽ�飺
����(1)�������x�{(di��o)�ء��~�G�نΰ������н����N�ЙC(j��)������(��(li��n)����ԭ�ͺͶ�h(hu��n)���N)�������������l(w��i)asI/lasR��rhlI/rhlRȺ��ϵ�y(t��ng)�����У���هRhl��QS�ɿ����S���N�ķֽ���x������_(d��)��
�������~�G�نΰ����У�ɫ���ή���^(gu��)�������������������ᣬ���ͨ�^(gu��)��Zͪ(PQS)����ϳ�;��ֱ�Ӽ���نΰ�����Zͪ��̖(h��o)����ͨ�^(gu��)ant-cat;���D(zhu��n)���������a(ch��n)������?y��)?j��)��(b��o)����������(y��ng)���T��(d��o)���ٲݿ�(�r(n��ng)ˎ���ЙC(j��)��Ⱦ��)��¶�£����c�U��������߀ԭ���oø��(NADPH)�a(ch��n)����ͬ�r(sh��)�p��߀ԭ���oø��(NADH)�Įa(ch��n)����
����(2)���{(di��o)���x�����D(zhu��n)䛡�Ⱥ�о�RshUS6-1��rsh�����mȻؓ(f��)�{(di��o)�ض�h(hu��n)���N�������P(gu��n)�Ļ�����_(d��)�����ɴ��M(j��n)��(x��)���m��(y��ng)�h(hu��n)����������������������������g�����(x��)�������h(hu��n)���N��Ч�ʡ�
����(3)⧜���̖(h��o)���ӡ��ֽ���x;����ijЩø�����g���x����F(xi��n)��Ⱥ�w⧜焩����⧜���̖(h��o)���Ӽ�����Ⱥ�w�Б�(y��ng)���緼�N�����^(gu��)���У�Hod�h(hu��n)�ѽ�ø�����ѽ�PQS��
����(4)ȫ���{(di��o)�ء���BkdR��֧�ͪ��Ó��ø���D(zhu��n)䛼��������̼ˮ������ֽ���xø��Ӱ푾Gē�غϳɣ��{(di��o)��(ji��)��Zͪ��̖(h��o)(PQS)QSϵ�y(t��ng)��ͨ�^(gu��)��������ጷż�(x��)����DNA�����㻯�����{���£�ȫ���{(di��o)��(ji��)��crc��ͨ�^(gu��)BkdR���Ƽ�(x��)����(n��i)̼ˮ������ֽ⡣
����4.5.2 ��(f��)�sȺ���wϵ
������(f��)�sϵ�y(t��ng)�������漰��(x��)���g������á���������Ⱥ��Y(ji��)��(g��u)���܌�(du��)AHLs�����У�����ͨ�^(gu��)�{(di��o)��QS��(y��u)��Ⱥ��Y(ji��)��(g��u)��������ꐴ��x;���̓�(y��u)�������a(ch��n)�������ģ����M(j��n)��Դ�ЙC(j��)��Ⱦ������オ��͵V����
�����O�ٔ�(sh��)��r�£���һ��(x��)�������h(hu��n)�������¿����M(j��n)���@�ý�����Ⱦ������д��x;������نΰ���Pseudomonas ADP���н��Ⱒ����������л���
������ϷNȺ�У�QS����ʹij��(g��)�NȺ��(du��)�����NȺ���Ѓ�(y��u)��(sh��)�����c���и�(j��ng)��(zh��ng)�P(gu��n)ϵ�ͷ����C(j��)�Ƶļ�(x��)������r(sh��)����QS�ļ�(x��)������ʹ��(j��ng)��(zh��ng)�Լ�(x��)��ֹͣ���߾������L(zh��ng)��
������������w����������Դ�����ɶ�NQS��������ȡ��AHLs���Һ���T��(d��o)����AHLsጷţ����M(j��n)��ΰ�����(Aeromonas)�ͼنΰ�����(Pseudomonas)�����������Ⱦ��ȥ��Ч����
����A. Valle��ͨ�^(gu��)�������������Դ����AHLs(3O-C6-HSL��C6-HSL)�����˱��ӽ��⣬���о��������Ͼ���(Thauera)�ă�(y��u)��(sh��)����(���ӽM)�������ΰ�����(Comomonas)(����+AHLs�M)ȡ������AHLs�錧(d��o)������Ⱥ��׃����
����δ��(l��i)��Ό��һ�M�b����V����Դ�ЙC(j��)���Ⱥ�w�Б�(y��ng)����M�ϣ���ֹ�ж����g�a(ch��n)��ķe����Ҫ�M(j��n)һ���о���
����05
����չ��
�����S���F(xi��n)���M�W(xu��)�ȷ����ֶεđ�(y��ng)�ã��ЙC(j��)��Ⱦ����⽵�⼰�{(di��o)�ؙC(j��)���о��õ��M(j��n)һ����չ��������EPS�M�ɳɷֺͿ��g�Y(ji��)��(g��u)��(f��)�s���Һ��в����ܽ����|(zh��)��δ��(l��i)EPS����������������W(xu��)�������д����ơ�
�������⣬�ЙC(j��)�オ���^(gu��)���еĻ��W(xu��)څ�����D(zhu��n)��{(di��o)�غ�Ⱥ�w�Б�(y��ng)�{(di��o)�ؙC(j��)�����д������о���δ��(l��i)�о�څ��(sh��)���Y(ji��)���£�
����(1)�R(sh��)�eEPS�c�ЙC(j��)��Ⱦ�����õ��P(gu��n)�I�M�֣�ͨ�^(gu��)����������ػ��������Á�(l��i)�{(di��o)��(ji��)����P(gu��n)�I�M�֣�����EPSȥ���ЙC(j��)���Ч��;����ģʽ������ģ�MEPS�ɷ֣���(g��u)���ЙC(j��)����xģ�ͣ��������挍(sh��)�ďUˮ̎��ϵ�y(t��ng)�(y��n)�C����Ч�ԡ�
����(2)�P(gu��n)עQS��(du��)����ȥ����Ⱦ����{(di��o)�ؙC(j��)�������û���M�W(xu��)���D(zhu��n)䛽M�W(xu��)�����x�M�W(xu��)�Ȱl(f��)��QS�{(di��o)��EPS�ϳɵĻ����w�M�֣�ͬ�r(sh��)�Y(ji��)�Ϲ��V���g(sh��)�z�y(c��)�{(di��o)�غ�EPS�c��Ⱦ��֮�g�����û��F(tu��n)��׃�����龫��(zh��n)QS�{(di��o)�؏Uˮ����̎�����ЙC(j��)��Ⱦ�オ���ṩ��Ч;����
����(3)�о���(f��)�sϵ�y(t��ng)�Б�(y��ng)��QS�{(di��o)����Ⱦ�オ���Ⱥ�䘋(g��u)���C(j��)������(f��)�sϵ�y(t��ng)��Ⱥ��M�ɵ�׃�����|(zh��)����ˮƽ�D(zhu��n)�ƣ�QS����QQ���Ͳ��a(ch��n)����푑�(y��ng)��̖(h��o)���ӵľ����ཻ������ʹ����Ⱦ��ȥ���C(j��)�Ƹ���(f��)�s����Ό��һ�M�b����V����Դ�ЙC(j��)���Ⱥ�w�Б�(y��ng)����M�ϣ���ֹ�ж����g�a(ch��n)��ķe�����д��M(j��n)һ���о����䌢��(du��)��(sh��)�H�Uˮ̎�����ЙC(j��)���ȥ��������Ҫ���x��
�w�S������1�����a(ch��n)�S��Ԕ���c(di��n)����http://m.quxianping.cn/



